Excavating the time frame of human evolution from its Y chromosome DNA
Next generation sequencing is now providing a flood of Y sequencing data. Appropriate software was developed to analyze these data and deduce the structure of the tree, finding new nodes and each new sequencing is providing a detailed branch of the tree. The part of the tree shown below is from an analysis by Yfull (www.Yfull.com – with permission) and the part shown is shared by all people of the large R1b-M269 group , about half of the men in western Europe countries such as France or England.
My aim with this article is to show that these trees with a large amount of SNPs can provide a frame to read the past, the different stages from the near origin of Homo sapiens to recent times. I call near origin the practical root of these trees. As shortly presented the very top of the tree has been subject to recent changes but we don't need these details ; the split from the A african group is close enough for the purpose of this article. As always on this blog I'll detail the reasoning allowing deduction on the history of the human group behind the SNP data, advanced readers will find some discussion on the possible impact of the mount Toba eruption on the human expansion. Surprisingly no team has published, so far, their own interpretation. Mine might be contested but it can also be a start for a better understanding ; this is my hope.
R1b-M269 branch of the Y tree
Y-root
A1 V250 L985 L1013 L1002 M9072 P305 V174 V161 • V161.1 • V161.2 L1005 L1112 V241 L986 L1004 L1084 V168 L989 L1009 Y1459 V171 V238 L1053
A1b P108 V221
BT V2952 • M9137 M9159 • PF767 M9380 • PF1256 M9312 M9382 • PF1257 V2167 • M9026 M9178 M9251 • PF913 M9133 • PF715 M9113 M8952 M9232 • PF880 M8961 • PF201 M9202 V3063 • M9145 • PF733 M8972 M9221 M299 M9356 M9070 M8968 • PF207 M8967 V2804 • M9124 • PF701 M9157 • PF766 M9081 M9378 M9379 • PF1253 V2561 • M9110 • PF684 M9054 M9361 M9349 M9036 • PF308 M9396 M9214 M9365 • PF1218 M9287 M9226 • PF869 M9057 M9389 M9359 M9338 • PF1064 M9046 • PF324 M9322 • PF1049 V2319 • M9031 M9027 L970 • PF1065 M9346 M9238 M9203 • PF837 M9240 • PF896 M9123 L957 M9305 • PF1022 M9372 V4201 • M9297 • PF1003 M9103 • PF679 M9292 • PF995 M9249 M9406 M9375 M9321 • PF1045 M8958 • PF196 M9278 • PF969 M9188 V1395.1 • M8997 • PF260 M9099 • PF674 M9104 • PF680 M9021 • PF288 V3546 • M9265 M9293 • PF997 V2579 • M9114 M9002 • PF267 M9409 • CTS12197 • PF1314 L978 • PF93 M8986 M9177 M9369 M9045 M9086 • PF648 V1158 • M8954 M9155 • PF762 M9328 • PF1053 M9310 M9127 M9399 • PF1283 M9329 V2634 • M9117 M8980 • PF229 M9174 M9373 M9311 • PF1030 M9198 M9242 • PF899 M9204 M9231 • PF876 Y1547_2 M9197 M9254 M9230 • PF870 M9102 M9173 • PF794 M9116 • PF688 V1561 • M9009 M9135 M9394 • PF1271 M9377 • PF1241 M9064 • PF350 M9246 M9398 • PF1279 V2209 • M9028 • PF298 M9410 M9083 M9196 M9098 M9097 • PF672 M9397 M9271 M9341 • PF1072 L969 M9213 V1530 • M9008 M9366 M9326 M9041 • PF319 M9316 • PF1034 V4025 • M9291 M9252 V2507 • M9038 • PF313 V235 • PF1410 PF1407 • V21 • M8969 M9069 • PF635 M9187 M9411 • PF1315 L1060 • PF1021 M9412 M9200 • PF835 M9354 M9223 • PF865 M9049 M9227 M9010 V1813 • M9019 • PF286 M9340 M9237 • PF890 V4213 • M9298 M9129 • PF707 M9172 M9220 V3107 • M9148 • PF744 M8951 V3636 • M9269 PAGE65.1 • SRY1532.1 • SRY10831.1 • SRY10831 • PAGE65.2 • SRY1532.2 • SRY10831.2 V2437 • M9034 V3226 • M9152 M9357 • PF1209 M9138 V1347 • M8994 P97 M9141 L1071 • M8945 M9136 • PF724 V2821 • M9126 • PF703 M9370 M9191 M9318 • PF1039 M9327 M9105 M8959 • PF198 M8983 • PF230 M9068 M9352 • PF1100 M9179 M9244 M9209 M9042 Y6870 PF1405 • V216 • M8953 PAGES00024 • M9160 M9343 • PF1084 M9334 M9003 V64 • PF1412 V2352 • L1062 • PF302 M9266 • PF946 M8976 • PF215 V202 • PF1404 M8985 • PF232 M9405 M9360 M9074 M8993 M9146 M9112 M9225 • PF868 V1730 • M9017 • PF282 M9193 L440 Y1546_2 V31 • L413 • PF1409 V29 • PF1408 M8971 M9302 V2397 • M9032 • PF304 M9393 M42 L438 M8955 • PF12 M9425 M9189 M9130 • PF708 M9107 M9015 V2000 • M9025 L418 M9075 M8960 • PF200 M9094 • PF671 M9219 • CTS7503 PAGES00026 • M9336 M9115 • PF687 M8988 M9290 • PF989 M9192 M9176 V3601 • M9267 • PF948 V3304 • M9257 V2318 • M9030 M9169 M9195 M9284 M9011 L971 M9420 M9331 • PF1057 M9217 • PF857 M9319 M9001 M9216 V3998 • M9288 • PF985 M8999 PF601 M9286 V3297 • M9255 • PF925 M9304 V3037 • M9143 • PF732 M9228 V3002 • M9140 M9039 M9353 M9280 L1061 • PF1101 M9077 M9020 • PF287 M9180 V3916 • M9283 • PF973 M8979 • PF226 M9315 • PF1033 M91 M9367 M9111 M9165 M9100 L977 M9199 • PF834 M9121 M9376 M9236 M9272 M9248 M9245 M9210 M9016 M9348 • PF1093 M9182 PF1406 • V102 M9215 • PF847 M9089 • PF653 V59 • PF1411 V187 • PF1403 M9317 M9260 M9235 • PF886 M9004 • PF270 M9125 M9043 M9056 M9344 M9368 M9325 • PF1052 M9066 M9087 L1220 • M9212 M9234 • PF885 M9285 M9296 V1456 • M9000 V2656 • PAGES00081 • M9118 M9131 V3347 • M9261 • PF931 M9390 • PF1262 V3032 • M9142 • PF731 M9166 • PF785 M9408 • PF1296 M8956 • PF14 M9335 • PF1060 M9306 M9323 • PF1050 M8977 M9006 M9404 M139 M9095 M9050 V4007 • M9289 • PF988 M94 • PF1081 M8973 • PF211 V3795 • M9277 M9303 M9301 • PF1015 M9258 M9175 V3357 • M9262 • PF932 M9417 M9128 M9224 L962 M9347 V1506 • M9005 V3904 • M9282 M9080 M9253 • PF914 V1015 • M8947 M9156 • PF764 M9400 • PF1284 M8957 V4130 • M9295 • PF1000 M9065 • PF351 L604 • PF1243 M9421 M9374 • Z4690 M9076 V2760 • PF699 • M251 • M9122 M9151 M9300 M9270 • PF952 M9139 M9163 • PF777 M9218 • PF860 M8970 • PF208 M9362 M9239 M9109 M9263
CT M5745 • CTS8608 M5708 Y1819_2 M5687 • CTS5019 Y1569 M5589 • PF212 PF1415 • V226 • M5603 Y1510 M5688 M5657 Y1544_1 M5751 • PF937 M5608 • PF258 M5821 • PF1269 M5651 • CTS1996 Y1441 M5743 • CTS8542 M168 • PF1416 M5588 • PF210 V2824 • M5671 • CTS3662 • PF704 M5593 M5810 • CTS11408 Y1508 M5699 • PF803 PF1016 M5753 • CTS9458 • PF947 Y1528 Y1581 Y1511 M5752 • CTS9296 V1863 • M9022 M5742 • PF904 Y1446 M5781 • PF1040 PF110 Y1575_1 Y1475 M5661 • CTS2842 M5786 • PF1061 Y1490 M5769 • PF996 Y1498 M5785 M5724 • PF866 M5777 Y1828 M5775 Y1482 M5609 M5718 • CTS7257 M5726 M5707 • CTS6383 Y1544_2 M5780 L1028 • CTS4368 • M5680 M5716 • PF840 M5759 M5725 • CTS7741 • PF867 Y1485 Y1507 M5738 M5798 PF1418 • V52 • M5721 M5732 • CTS8089 Y1503 M5660 Y1531 V3648 • M5760 • PF954 M5811 • PF1238 Y1571 M5613 M5692 M5683 • CTS4650 M5805 • PF1227 M5768 Y1489 V1494 • M5615 • PF269 Y1817 Y1509 M5584 • CTS543 • PF206 M5715 • CTS6907 • PF833 PF192 Y1505 M5670 • CTS3460 Y1452 M5587 Y1579 M5819 M5597 • CTS1217 Y1448 M5803 M5747 Y1471 M5600 Y1455 V1052 • M5576 • CTS125 Y1496 Y1460 V3623 • M5754 M5686 Y1546_1 Y1585 M5647 Y1465 M5717 • PF844 M5825 M5791 • PF1080 Y1464 M5617 • PF274 Y1574 M5630 M5614 • PF266 M5629 V3641 • M5757 • CTS9555 Y1494 Y1567 M5648 Y1594 Y1578 M5817 M5830 • CTS12633 • PF1329 M5606 • PF256 Y1450 M5831 Y1488 M5795 M5813 M5712 Y1504 M5772 • CTS10512 V3642 • M5758 • CTS9556 V3310 • M5748 • CTS8980 • PF928 M5801 V4106 • CTS10362 • PF998 • M5770 M5607 M5808 • CTS11358 M5665 • CTS3216 M5636 Y1457 Y1524 M5784 • PF1059 Y1474 M5656 M5621 M5616 • PF272 V1431 • M5612 Y1492 M5720 • CTS7482 Y1480 Y1521 M5746 • CTS8709 M5578 M5767 • CTS10110 Y1527 M5641 M5796 • PF1097 V3337 • M5750 • CTS9014 Y1470 M5832 • PF1333 M5723 • PF862 CTS5457 M5741 M5653 • CTS2077 • PF657 M5590 • PF216 M5598 M5697 • CTS5746 M5800 • PF1203 Y1589 PF1413 • V189 • M5577 M5762 • CTS9722 Y1568 M5705 • CTS6327 • PF811 PF500 Y1506 Y1593 M5809 • PF1237 V1043 • CTS109 • M8948 Y1552_2 M5713 • CTS6800 Y1502 M5618 Y1538 V2216 • M5633 M294 M5776 • PF1029 V3858 • PF970 M5794 • PF1092 M5711 • PF821 Y1587 M5602 • PF246 Y1467 M5802 V2901 • M5675 • PF719 M5783 V3908 • M5766 V1325 • M5605 M5591 • PF223 Y1497 Y1599 Y1819_1 Y1473 M5816 • CTS11827 M5698 • PF796 Y1525 Y1559_2 M9150 • PF750 M5797 • PF1098 M5649 PF342 Y1454 Y1438 Y1447 V2175 • M5632 M5709 M5662 PF1414 • V9 • M5585 V3758 • M5764 • CTS9828 • PF964 Y1491 Y1476 M5601 Y1472 M5765 M5628 Y1495 M5611 • PF263 Y1461 M5689 M5626 PF1276 M5700 • CTS6252 M5642 V4162 • M5771 M5599 • PF234 Y1456 M5722 • CTS7517 M5594 • CTS1109 Y1443 M5582 • CTS401 • PF202 M5736 • CTS8243 • PF891 M5730 Y1586 Y1573 Y1518 M5814 M5788 V3808 • Y1539 M5739 • PF898 PF1417 • V41 • M5695 Y1591 M5659 • PF667 M5645 M5729 • CTS7936 M5691 • PF779 M5822 M5664 • CTS3120 • PF683 Y1462 V3317 • M5749 M5778 • PF1031 M5681 PF1420 • V55 V1401 • M5610 M5650 V1653 • M5625 Y1440 Y1791 M5737 • PF892 Y1580 Y1590 M5706 • PF815 Y1577 M5792 • PF1088 M5678 • PF725 Y1514 Y1559_1 M5622 Y1469 M5826 M5620 CTS11575 • PF1245 M5583 • CTS423 V3728.1 • M5763 • CTS9760 • PF961 M5640 • PF318 M5714 • CTS6890 M5790 V1878 • M5631 • PF292 M5719 • PF850 M5652 • PF652 Y1483 M5627 Y1451 M5639 Y1444 M5684 • CTS4740 • PF751 M5782 • PF1046 M5694 • CTS5532 Y1458 M5735 • CTS8166 M5690 • CTS5318 Y1449 M5679 • CTS4364 M5638 • PF316 M5624 P9.1 • P9 M5728 • CTS7933 M5676 • PF720 Y1526 M5818 • CTS11991 M5682 M5812 V1540 • M5619 • PF278 PF15 M5669 • CTS3431 M5727 • CTS7922 • PF875 M5595 • CTS1181 M5774 M5804 • CTS10946 Y1537 M5756 • PF951 M5646
CF M3711 • CTS6376 • PF2697 P143 • PF2587 CTS3818 • PF2668 • M3690 V3489 • PF2723 • M3727 • F2841
F PF2651 • F1704 • M3675 Y1812 L498 • PF2707 • M3717 CTS4969 • PF2682 • M3700 PF2743 • M3747 Y1810 PF2615 • M3652 PF2756 CTS11819 • PF2766 • M3761 M213 • P137 • PF2673 • PAGES00038 P316 • PF2696 M3771 • CTS12632 • PF2775 M3693 • CTS4139 • PF2672 YSC0001298 • PF2620 • F1302 • M3656 Y1805 PF2744 • M3748 PF2653 • F1714 • M3677 P138 • PF2655 PF2639 M3696 • CTS4443 • PF2677 PF2611 • M3647 PF2608 PF2660 • F1767 • M3683 M3692 • CTS3996 • PF2671 L468 • PF2689 • M3703 PF2588 • M3635 • CTS71 M3640 • CTS540 • PF1506 CTS2041 • PF2652 • M3676 M3687 • CTS3195 • PF2664 L929 • PF2605 • M3643 PF2740 • M3744 CTS3536 • PF2666 • M3688 V1644 • L313 • PF1426 • M3651 PF2739 • M3743 P151 • PF2625 M3728 • CTS9280 • PF2724 M3756 • CTS10983 • PF2760 CTS7981 • PF2710 PF2601 • M3641 • CTS608 M3720 • CTS8014 • PF2711 CTS5432 • PF2687 • M3702 PF2592 PF2637 • M3672 P158 • PF2706 CTS1468 • PF2607 • M3644 PF2742 • F3254 • M3746 L882 • PF2745 • M3749 CTS10290 • PF2735 • M3739 M3730 • CTS9372 • PF2725 CTS11726 • PF2765 P166 • PF2702 CTS12138 • PF2774 • M3770 PF2718 • F2710 • M3723 PF2729 • F2964 V3940 • PF2732 • F2993 • M3737 M3706 • CTS5948 • PF1695 L352 • PF2728 • M3734 F3561 • M3766 Y1808 PF2630 • M3664 PF2593 P161 • PF2719 CTS3868 • PF2669 PF2700 • F2402 • M3714 PF2737 • F3136 • M3741 P163 • PF2686 M3729 • CTS9317 • PF1767 M3689 • CTS3654 • PF2667 CTS12027 • PF2768 • M3763 P160 • PF2618 Y1799 Y1807 P14 • PF2704 L1074 • CTS4267 • PF2674 • M3694 L851 • CTS11821 • PF2767 • M3762 PF2647 P140 • PF2703 PF2612 • M3648 M3718 • CTS7878 F3556 • M3765 • PF1914 Y1809 CTS4557 • PF2679 • M3698 PF2629 • M3663 V3268 • PF2683 • F2048 • M3701 PF2747 • M3750 PF2688 • F2142 V2513 • L543 • PF2663 • M3686 P159 • PF2717 PF2609 • M3645 P135 • PF2741 CTS11150 • PF2761 • M3758 P157 • PF2771 CTS1932 • PF2650 • M3674 CTS6135 • PF2693 • M3708 PF2736 • F3111 • M3740 M235 • PF2665 • PAGE80 P187 • PF2632 PF2627 • M3661 P139 • PF2698 M3732 • CTS9456 • PF1438 M3731 • CTS9418 • PF2726 M3712 • CTS6542 • PF2699 PF2594 V1990 • YSC0001297 • F1209 • M3654 M3725 • CTS8985 • PF2721 L350 • PF2692 • M3707 CTS9534 • PF2727 • M3733 PF2750 • M3753 Y1811 V3919 • PF2731 • F2985 • M3736 Y1802 Y1758 M3699 • CTS4737 • PF2680 F3512 • PF1911 P148 • PF2734 Y1820 PF2628 • M3662 PF2590 • V205 • M3638 F3584 • M3768 • PF1916 Y2888 F773 P149 • PF2720 V1597 • PF2614 • F1089 • M3649 M89 • PF2746 CTS11370 • PF2763 • M3759 V3900 • L470 • PF2730 • M3735 PF2591 • M3639 M3721 • CTS8467 • PF2715 P133 • PF2636 Y4457 PF2589 • V186 • M3637 V1029 • F719 • M3636 Y1800 PF2600 CTS2097 • PF2654 • M3678 PF2631 • M3665 F3692 • M3650 PF2713 PF2598 P136 • PF2762 L132.1 • L132 • L132.2 • PF1437 M3713 • CTS6843 • PF1720 CTS5750 • PF2691 • M3705 V1355 • YSC0001295 • PF2610 • F1046 • M3646 M3716 • CTS7301 • PF2705 P142 • PF2604 CTS7002 • PF2701 • M3715 PF2613 CTS5264 • PF2684 V2194 • PF2619 • F1285 • M3655 P141 • PF2602 M3724 • CTS8638 PF2621 • F1320 • M3657 PF2758 • F3335 • M3754 P146 • PF2623 PF2772 • F3616 • M3769 P134 • PF2606 M3666 • PF1580 P145 • PF2617 PF2749 • M3752 PF2616 • F1149 • M3653 Y1813 M3682 • CTS2480 • PF2659
GHIJK M3684 • CTS2569 • PF2661 M3680 • CTS2254 • PF2657 M3773 • CTS12673 V2308 • YSC0001299 • PF2622 • F1329 • M3658
HIJK PF3494 • F929 • M578
IJK L16 • M522 • S138 • PF3493 V1438 • YSC0001319 • PF3497 • M2684 L15 • M523 • S137 • PF3492 • Z4413 PF3500 • M2696 V1295 • PF3495 • F3689 • M2682 • Y2571
K P132 • PF5480 PF5500 • F2548 • M2692 L819 • CTS4265 • PF5494 • M2686 V3169 • PF5495 • F2006 • M2688 PF5470 M2352 • CTS2071 • PF5489 YSC0000055 • PF5459 • M2348 P131 • PF5493 M9 • PF5506 L469 • PF5499 • M2689 PF5469 • V104 PF5488 • M2351 V4038 • PF5503 • F3026 • M2694 P128 • PF5504 YSC0000222 • PF5505 • L1346 • M2695 CTS9278 • PF5501 • M2693 CTS10976 • PF5509 • M2698
K(xLT) M526 • PF5979
MP-M1205 PF5852 PF5969 L405 • PF5990 P331 • M1221 • YSC0000186 • PF5911 M1205 • CTS2019
P P237 • PF5873 CTS12028 • PF5977 • M1272 F313 • M1219 YSC0001257 • CTS1907 • PF5894 • M1204 M1235 • CTS7604 • PF5928 M45 • PF5962 L741 • PF5960 • YSC0000297 PF5849 Y503_1 M1270 • CTS11173 • PF5974 M1149 • CTS10168 • PF6061 P235 • PF5946 PF5878 • M1194 PF5985 PF5945 • F524 • M1248 PF5991 • F4 • M1183 M1228 • CTS5884 • PF5917 V1809.1 • PF5872 • M1192 CTS3316 • M1209 CTS5808 • PF5915 • M1226 Y451 PF5854 PF5914 • F332 • M1224 Y272 M1264 • YSC0000227 PF5483 PF5871 • M1190 92R7_1 PF5886 • M1199 P228 • PF5927 Y44 • M1207 M1244 • CTS9604 PF5975 • F640 • M1271 CTS6948 • PF5922 • M1231 V2974 • L779 • PF5907 • YSC0000251 V1195 • PF5861 • F83 • M1185 YSC0001285 • CTS5673 • PF5497 • M1225 P230 • PF5925 CTS3775 • PF5906 • M1214 L768 • PF5976 • YSC0000274 PF5848 PF5964 • M1263 P283 • PF5966 PF5958 • M1160 P226 • PF5879 P295 • S8 • PF5866 CTS3736 • PF5905 • M1213 PF5892 • M1202 CTS1518 PF5876 • M1193 PF5901 • F1857 • PAGES00083 • PAGE83 PF5951 • F556 Y448 PF5461 PF5993 • Z1244 V231 • PF5862 • F91 PF5891 PF5881 • F180 • M1196 Y444 PF5956 • M1259 Y45 • M1208 CTS3697 • PF5904 • M1212 M1186 • YSC0000279 • PF5864 V2979 • M1216 • YSC0000176 • PF5908 V1651 • PF5870 • F115 • M1189 P243 • PF5874 L82 • PF5972 Y503_2 PF5994 V3529 • F506 • PF5940 • M1243 • YSC0000966 M1240 • YSC0000205 • PF5936 PF5916 • F344 • M1227 L721 • PF6020 PF5887 V3732 • M1246 • YSC0000270 • PF5943 PF5882 CTS5418 • PF5912 • M1222 Y455 PF5846 • M1184 • CTS216 V1079 • PF5845 • CTS196 PF5954 • M1256 PF5970 Y269_1 PF5984 • F680 • M1275 PF5944 • F521 • M1247 P282 • PF5932 PF5957 • M1260 PF5867 CTS7886 • PF5929 • M1236 Y450 V3240 • M1218 • CTS4944 • PF5909 P284 L781 • PF5875 • YSC0000255 PF5850 PF5888 P244 • PF5896 • P244.1 • P244.2 P240 • PF5897 L1185 • CTS9162 • PF5937 • M1241 PF5869 • M1188 PF5971 PF5978 • F647 • M1273 CTS3358 • M1210 • PF5899 Y456 CTS12299 • PF5987 P69 Y447 PF5885 • F212 • M1198 PF5883 • M1197 L268 • PF5983 P27.1_1 • P207 • P27.2_1 PF5853 L138 PF5855 PF5880 • M1195 Y446 V4004 • PF5949 • F536 • M1251 Y1403 M1269 • CTS10859 P281 • PF5941 PF5980 • F653 Y458 Y483_1 M74 • N12 • PF5963 PF5851 L536 • PF5860 L471 • PF5989 P239 • PF5930 PF5955 • M1257 PF5981 • F671 • M1274 PF5982 L821 • PF5857 • F29 • M5579 PF5471 PF5473 M1250 • CTS10085 • PF5948 M1109 • CTS4437 CTS3813 • PF5491 • M1215 PF5865 • M1187
R M732 • CTS8311 • PF6055 CTS3622 • PF6037 YSC0000201 • PF6057 • M734 • S4 P227 P280 • PF6068 Y453 M696 • CTS5815 • PF6044 P232 P224 • PF6050 YSC0000232 • M789 • L1225 • PF6076 P285 • PF6059 CTS2913 • PF6034 • M667 V2573 • YSC0001265 • CTS3229 • PF6036 • M672 PF5953 • M764 M207 • UTY2 • PF6038 • PAGES00037 V3466 • CTS9200 • PF5938 Y457 YSC0000233 • PF6077 • L1347 • M792 Y480 P229 • PF6019 Y472 • F47 • M607 • PF6014 • S9 M795 • CTS11075 • PF6078
R-Y482 M799 • PF6079 • YSC237 PF6040 • YSC179 PF5919 • F356 • M703 Y482 • PF6056 • F459
R1 M813 • CTS12618 • PF6089 L875 • PF6131 • YSC0000288 • M706 M730 • CTS8116 • PF6138 P231 P236 • PF6137 M717 • CTS7122 • PF6135 P245 • PF6117 P233 • PF6142 PF6146 M173 • P241 • PF6126 • PAGES00029 Y464 • PF6008 PF6069 M781 • PF6145 PF6073 P286 • PF6136 CTS3321 • PF6125 • M673 CTS2680 Y305 • PF6031 Y512 Y290 • F211 PF6118 • M640 M812 • CTS12546 • PF6088 PF6110 PF6011 M663 • CTS2565 • PF6122 V1478 • PF6116 • F102 • M625 P234 • PF6141 YSC0000230 • L1352 • M785 PF6133 • F378 • M711 Y465 V1356 • PF6114 • F93 • M621 P242 • PF6113 P294 • PF6112 F132 • M632 M643 PF6119 M748 • YSC0000207 PF5477 • F28 CTS3123 • PF6124 • M670 M306 • S1 • PF6147 Y459 P225 • PF6128 P238 • PF6115 M714 • CTS7066 • PF6049
R1b M343 • PF6242
R1b1 L278 M415 • PF6251
R-L389 L389 • PF6531
R1b1a L320 • PF6092 P297 • PF6398
R1b1a2 PF6410 • M520 L265 • PF6431 M269 • PF6517 YSC0000269 • PF6475 • S17 S3 • PF6485 S10 • PF6399 L1063 • CTS8728 • PF6480 • S13
Each « code » as P108 is the name of a SNP (see A1b for P108 - at the top of this tree), it's a mutation on the Y chromosome and let's recall that these mutations are all neutral, meaning that they are not associated with any phenotype, no visible changes. They are merely DNA changes detected by sequencing (no other way) and used as markers. As there are several research teams and each is giving their own names, one single mutation can get several names and this tree is accounting for this fact with the following mark : [ • ] , separating the different names for one same mutation. For example L1063 • CTS8728 • PF6480 • S13 (just above, see bottom of this tree) is showing 4 different names equivalent for what is 1 SNP and counted as such in the counting to come.
A sequencing is providing more SNPs than shown here as « recent » SNPs at the bottom of the tree, below M269 were removed. The reason for this is that the bottom of the tree is bushy and still difficult to read ; this article is only focusing on the history « available » between A1b / [BT] and M269. In France (the same in England) about half of the men have this same segment of the Y tree. From the top of the tree (as displayed here) to M269 the SNP count is roughly 1000. It's important to understand that the DNA sequencing producing the data (mostly the « Big Y » from FTDNA company) is far from finding all SNPs : the depth of the sequencing can be improve and it would allow detecting more SNPs. These data are usable (my opinion) as they reflect a percentage of the total, already in significant counts.
The early Homo sapiens fossils are pointing to the time range between 160 000 – 180 000 years ago (with still a margin of error). I took 170 000 years ago for the time range and the SNP count was estimated around 1200 when considering mean branch length till today (not limited to M269) corresponding to an average time interval between 2 SNPs of 140 years. On this basis we can read the tree presented above.
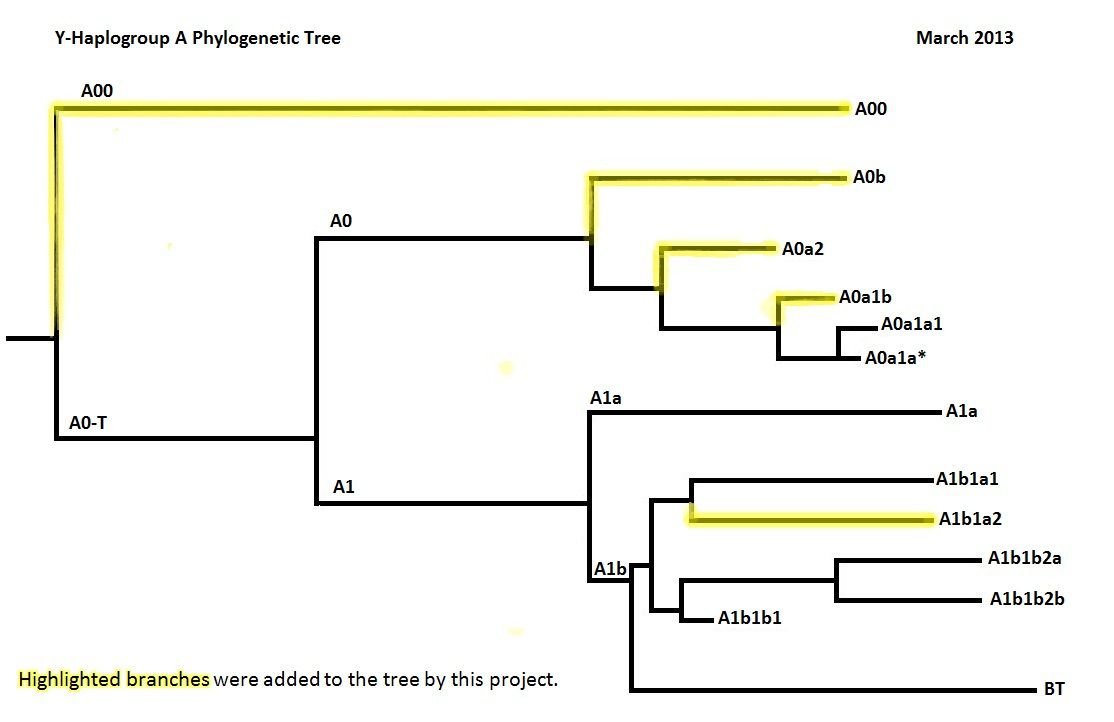
The top of the Y tree was found to be far more complex with the discovery of rare variants ; below is the root of the Y tree as published in 2013. I won't use it but I wanted to account for it :
• A00 AF4, AF5, AF6/L1284, AF7, AF8, AF9, AF10, AF13, L1086, L1087, L1088, L1091, L1092, L1094, L1096, L1097, L1102, L1103, L1104, L1106, L1107, L1108, L1109, L1110, L1111, L1113, L1114, L1115, L1117, L1119, L1122, L1126, L1131, L1133, L1134, L1138, L1139, L1140, L1141, L1144, L1146, L1147, L1148, L1151, L1152, L1154, L1156, L1157, L1158, L1159, L1160, L1161, L1163, L1233, L1234, L1236
• A0-T AF3, L1085, L1089, L1090, L1093, L1095, L1098, L1099, L1101, L1105, L1116, L1118, L1120, L1121, L1123, L1124, L1125, L1127, L1128, L1129, L1130, L1132, L1135, L1136, L1137, L1142, L1143, L1145, L1150, L1155, L1235
• • A0 L529.2, L896, L982, L984, L990, CTS2809/L991, L993, L995, L997, L998, L999, L1000, L1001, L1006, L1008, L1010, L1012, L1016, L1018, L1055, V148, V149, V154, V164, V165, V166, V167, V172, V173, V176, V177, V190, V196, V223, V225, V229, V233, V239
• • A1 L985, L986, L989, L1002, L1003, L1004, L1005, L1009, L1013, L1053, L1084, L1112, L1153, P305, V161.2, V168, V171, V174, V238, V241, V250
• • • A1* -
• • • A1a M31, P82, V4, V14, V15, V25, V26, V28, V30, V40, V48, V57, V58, V63, V191, V201, V204, V215
• • • A1b P108, V221
• • • • A1b* -
• • • • A1b1 L419/PF712
I just wanted to present the the deep rooting among A haplogroups, all being african while I'll using a simplified connection between A1b and [BT] (see below). So, just like fossils show it there must have been an origin of Homo sapiens in Africa. Again, I am not showing it but it would be a wrong idea that people, like Bushmen people, belonging to A0 groups (for some of them) have a less complex history ; there is a branch going to these A0 men with just as many SNPs than shown here for the R1b branch.
Below is another way to picture the complexity of the top of the Y tree and this is showing the BT branch as one outcome among many, all are localized in Africa with the only exception of the BT branch.

[BT] produced groups localized in Africa as are all B branch groups ; B groups are found among Pygmies, for example and [BT] produced all groups outside Africa (see below).
We now start the analysis of the SNPs and the first point is about the way SNPs come together in dense packs of SNPs : a node. The [BT] node displayed here is 353 SNPs large (see the tree of SNP above – in blue). Again, more SNPs will be found (more are already known) but thanks to the coherent displaying of the same percentage because most results are from sequencing by Big Y test (fro FTDNA company) that comparison between nodes will be possible. Trees with nearly all SNPs will come in a few years from now.
Now, let's think about it : what do these 353 SNPs at this « node » can mean ? 353 with a time interval of 140 years (in average) between 2 SNPs is a period of time 50 000 years long. So, it's from 170 000 years ago to 120 000 years ago in the last interglacial (LIG) usually said between -130 000 and -115 000. Here is a measurement of oxygen 18 in ice correlating with temperatures :

It can't be by chance that the end of the node correlates with cooling temperatures. It's easy to see also that the temperatures were cold until -130 000 and one may wonder why one same group is overlapping the cold and warm period. It could be that a branch was lost (in fact many branches were lost – I meant a main branching like those produced by the last glaciation – R1a / R1b). Impossible to tell where the [BT] people were localized but [BT] is splitting in [CT] (discussed below) and B , a branch found among african groups like Pygmies ; a localization in Africa is still likely but B groups going back to Africa (because of temperature change) is a possibility if [BT] was localized close enough ; old remains of modern humans were found in Palestine / Israel, a possible place for [BT] between -130 000 and -120 000 (of course my dating of -120 000 with the margin of error can be -115 000 – the end of the warm period).
Every SNP can be seen as a putative branching and the ancestral branch (not carrying the mutation) died out. This dilution out of so many branches is explained by a genetic drift and this in turn is only possible with a not too large group. A bottle neck, a period of time with a drastic decrease of population size is then likely at the bottom line of the [BT] node, -120 000 years ago when temperatures went down quickly. It's important to really catch this point : among the few lucky survivors one man had the succession of SNPs known today and a statistical effect made his descent to win over all others. Simulations show that this is what is expected when every man has only 2 surviving children in average : one boy and one girl ; hence any fluctuation (2 girls) is stopping a Y lineage and the result is : only 1 lineage left (for Y chromosome) with the specific set of SNPs now carried by all descent.
The small size of a group separated from other african groups might have allowed specific traits to evolve in the emerging new groups. As mentioned above the split 120 000 years ago separates the B group from a group, now called [CT] carrying the well known M168 marker, the first known of the set of now over 300 SNPs . The M168 marker is found among ALL ethnic groups out of Africa but not in those specific african groups like A0 and B groups already discussed. This finding was at the onset of the so called « out of Africa » theory. As discussed above it's likely that the BT group was already outside of Africa and the logic of the people proposing this theory wasn't strong enough. Also, the timing presented here is fairly different from the timing too often given with no data sustaining it.
I find it logical that the group who populated parts outside Africa when all other groups from the top of the Y tree are african might have lived already outside of Africa. Dienekes Pontikos was one of the first to propose the arabian peninsula as a place for such a stage and I find it a good idea as old human industries are well known there but this is not the only possible place. Northern area were still under Neandertal control and, as temperature was going down the limit might have left little room for modern humans. Indeed a near coastal stay (for example at the mouth of the Tigre river) would be in accordance with the next split in a coastal route of expansion and a land one, as discussed below.The SNP count is 319 making it (still with t=140) a 45000 years long period in round numbers. Hence, from 120 000 years ago to 75000 years ago the [CT] group developped in a changing environment with changing temperature.
Just like a bottleneck was implied for the end of the [BT] node, another bottleneck could explain the -75000 figure. As already discussed on this blog a giant eruption took place at mount Toba, dated -73000 with a margin of error of 4000 years. Scientists are still discussing the exact impact on the whole planet but the more it's studied the more the impact appears strong. It was sudden and temperatures dropped by 10 degrees, some say 15 degrees. Hence it can't be a coincidence if a major change is detected at that time. To better discuss the timing we need to look forward at what happened next . There was a split is [CF] and [DE] and this seems to correlate with the known split in the mitochondrial tree in N and M branches. The [DE] / M group has been associated with a coastal route, possibly on early rafts, a quick way to progress around India. The idea is that this expansion was started at the time of the eruption. Some |DE] might have been already at the Ganges delta while some might have populated the south border of the arabian peninsula and possibly the nearby horn of Africa. Why so ? We won't follow anymore the [DE] branch but the data are saying that [DE] was cut in 2 distant pools and these never met again. One pool of [DE] evolved in D, mainly in Japan today but it was in all south Asia and some are in Tibet (obviously a refuge) and one pool of [DE] evolved in E at the horn of Africa. Very rare [DE] cases were found south of Tibet and one should understand that it's a third branch (may be more) saying that once [DE] people not yet D or E settled there. I insisted on the explanation of the [DE] fate because it's part of the indication leading to the fact that Toba eruption had a deep impact and [CT] ends with the onset of 2 pools of [DE] with no more connection between them. [CT] also evolved in [CF] (see R1b-Tree in blue above) and these might have been less impacted as in a more remote localization from the Toba volcano.
The next C and F split occurs quickly after and one can see it as one group staying in place and one group (C) expanding in a novel environment. So, one word on the C branching as the R1b branch drives us on the F side. There are data suggesting that C haplogroup successfully went in all part of te world as a first wave, later displaced by others. Aboriginal Australian have the C4 branch, probably depicting the first men in Australia while the Mongol Genghis Kahn belong to the C5 branch. Some C people might have reached America but only rare cases are found today. Old remains of the C6 branch were recently found in Europe. So, except Africa, C men went everywhere it seems.
The F group is at the root of most groups populating Europe and Asia today as all neolithics are from this branch. SNPs count is 163 corresponding to 23000 years. Hence this large F node is from -75000 to -52000. The following steps are several branching which mean that rather than a bottle neck the « end » of the F node is when groups from the F « nest » started going in all directions, including Europe. Yet all these 163 SNPs mean that 163 times the ancestral branch was diluted out, meaning a very small group at some times. The temperatures as known by oxygen 18 concentrations in ice had a warming up around -60 000 , so this expansion is late relative to the improved conditions, possibly meaning a first phase of local expansion followed by dispersal in G, H, [IJ], and then L ; T and |MP]. At this time Neandertal starts retreating and modern man takes its place. I don't know of any accepted explanation for this change. Genetics and the time intervals found here are pointing for a significant progress in adaptation to cold of modern humans 60 000 years ago.
One word about the next step as it may give some clue on the positioning of the [MP] part of the original F group. The M haplogroup is mainly restricted to papuan in new Guinea, probably a refuge when neolithics populated south Asia but this can only be if [MP] was much closer to Indonesia than to the Indus basin. If M went south, P probably went in a more northern place just east of present day China to account for the M cousins. The 47000 years old Ust’-Ishim bones were tested of the parent group of P (K (xLT) and found to have large Neandertal proportion of DNA ; the expansion to northern areas might have been restricted to those who had the Neandertal DNA.
The P node is 135 SNPs large which fits with a 19000 years long period. Hence we are dealing with the time interval from -50000 to - 31000, a relative warming before the last glaciation. A bottleneck might explain the set of SNPs as before and it was followed by a split in R and Q groups. Q was clearly adapted to cold as this group managed to go through the Bering straight and get in America where it's (by far) the main haplogroup among amerindians. So, P, the parent group was a transition to a colder environment, possibly thanks to needles and clothing with instances known from 28000 years ago and it might be slightly older.

The diagram above is summarizing the fate of P group with a focus on R1a and R1b haplogroups. SNP counts are depicted in blue for each branch. I'll be using these counts. The ice age west of China isn't very well known but R node (40 SNPs) would end some 26000 years ago shortly before the ice maximum when temperatures dropped to a bottom level. The 24000 years old Mal'ta mammoth hunter encampment had bones tested of R haplogroup. This dating is with the margin of error and fits with this broad timing. This R group, in turn splits into a southern group R2 and the R1 group before the ice maximum and I would interpret it as a very difficult time for a group near extinction (so, again, a bottleneck). At ice maximum the R1 group splits into 2 subgroups seen as 2 geographical localizations that got separated and started evolving separately. These 2 localizations are not known but it should be clear that it's unlikely in Europe though there are indications that R1 was in Europe. 107 SNPs until R1b-M269 would make this important stage of the R1b branch only 6000 years ago but the precision isn't enough ; this is the limit of this approach and it's important to understand it not to say stupidities. Recent times were characterized by fast changes ; in the case of R1b-M269 found here to be 6000 years old a 3000 years change, well in the margin of error, is in accordance with more precise timings done with other methods. That's all.
In conclusion, the approach described here found a correlation between bottleneck of key stages and major climate changes. The description of the fate of the [DE] branch attempted here is providing a coherent explanation of the so called « out of Africa » expansion with a modified timing as explained. In turn the common ancestry in Africa while not denied is from a much deeper ancestry than it was said too many times. This approach will have to be refined but, as it is, it gives a time frame of early events.
[ Many thanks to the Yfull team for their SNP tree and permission to reproduce it. ]

























{kind=link}
{kind=link}